The Physical Rules
of Nutrition, Metabolism, and Exercise Science
Getting Rid of Junk/Damage
Reference source/supplementary reading for lecture Your Brain on Collision Sports
Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.4.3.4 Particle Clearance from the Lymphatics
Aside from the mucociliary escalator (Section 15.4.3.3.3), human biology includes no systematic mechanism by which foreign particles, once internalized in blood or tissues, can be physically ejected from the body.* Rather, physiological systems circulate particles through various “clearance” systems until one of two things happens: (1) the particles are chemically broken down, digested, or dissolved, with the remains subsequently metabolized or excreted (or for some fluorocarbons, exhaled as a vapor through the lungs [3127]), or (2) the particles are trapped somewhere in the body, more or less permanently. For insoluble, inert particles such as immobile diamondoid medical nanorobots, the first option is not available. Ultimately, these particles are either granulomatized in place or in bone (Section 15.4.3.5), or are swept into the lymphatic circulation and finally sequestered in the lymph nodes.
* Some phagocytic cells can transport small particles from elsewhere in the body to the lungs, from which it is then possible to extrude the particles through the lung wall and exhale them through the airway [2871]. There is at least one report of 0.5- to 5-micron alumina particles originally implanted subcutaneously and intraarticularly in mice that were later observed in the interstitium of the lung [1050], and similar observations have been reported for ingested 5.7-micron polystyrene particles internalized through Peyer’s patches [3047] and for Teflon particles [1392] injected into humans for the treatment of urinary incontinence [1277, 1280, 1286–1288] and vesicoureteric reflux [1312, 1401, 1403] that were subsequently transported to the lungs. However, while it is well known that particles which have crossed from the alveolar spaces into the lung interstitium can be ingested and returned to the alveolar spaces by arriving neutrophils or interstitial macrophages, the author has found no confirmation in the medical research literature that systematic long-range particle scavenging by migrating phagocytes followed by discharge in the lungs has been directly observed.
The lymphatic circulation collects foreign particles from all over the body – including tissue spaces such as the peritoneum [2998], dermis [3128, 3176], footpads [3129, 3130], and organs such as the liver [2670, 3131, 3132], spleen [2670], heart [3133] and lung [3134]. For example, metallic and polyethylene wear particles, mostly <1 micron in size, in patients with hip and knee replacement prostheses can migrate to liver, spleen, and abdominal paraaortic lymph nodes [2670]. Crystalline silica particles in a lipstick cream can enter the body through a recurring angular cheilitis (lip sore) and migrate to the submasseteric lymph node, forming a silica granuloma there [3176]. Inhaled silica particles can translocate from lung to hilar lymph nodes, enlarging the nodes with many granulomas containing silica and macrophages [3134]. Dodson et al [3135] suggest that lymph nodes “may be better indicators of lifetime exposure to dust than lung tissue” because the nodes permanently sequester indigestible particles like asbestos and retain this particle burden for the lifetime of the organism.
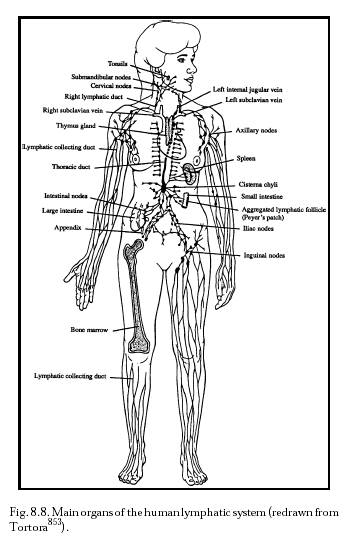
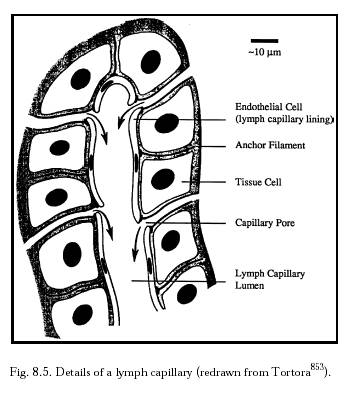
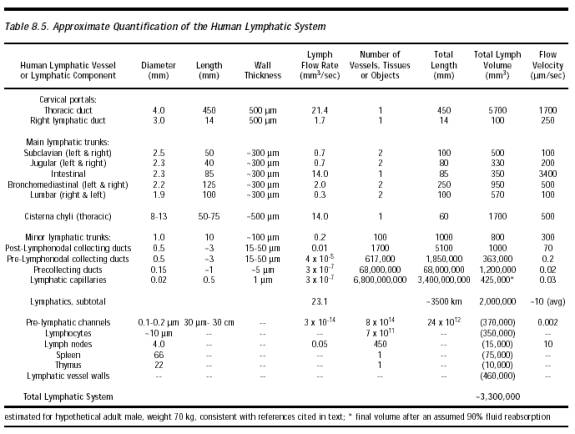
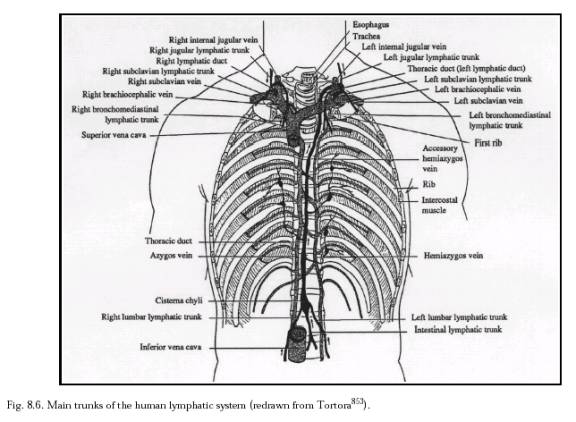
The lymphatic system (Section 8.2.1.3; Figure 8.8) is an auxiliary circulatory system in which interstitial fluid is drained off into tiny open-ended vessels, the lymphatic capillaries, whose walls are comprised of endothelial cells with a resting intercellular gap (Figure 8.5) normally ranging ~0.1 micron to several microns [4597]. However, Allen [4596] intraperitoneally injected particles up to 22.5 microns in diameter and all sizes later appeared in the diaphragmatic lymph. This suggested that the peritoneal mesothelium and the lymphatic endothelium on either side of the fenestrations of the basement membrane can open at least this wide to admit stray particles. Median lymph flow rate as measured by 1-micron latex microspheres in mouse tail skin lymphatic capillaries is 4.7 microns/sec, with pulsations synchronized with the murine respiration rate of 2 Hz [3136]. In supine humans [3137], the median resting flow velocity is 9.7 micron/sec in a 54.8-micron diameter lymphatic capillary, but this varies greatly with many factors including body orientation and vessel size (Table 8.5). The lymphatic capillaries gradually combine into larger and larger tubes until the largest are the size of veins (Table 8.5). Most lymph collects into the largest lymphatic of all, the thoracic duct, which leads into the left subclavian vein in the upper chest (Figure 8.6), thus returning the lymph to the blood circulation. (The lymphatics on the right side of the head, neck, heart, and thorax, and the right arm and lung, drain into the right lymphatic duct and thence to the right subclavian vein; see Figure 8.7.)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
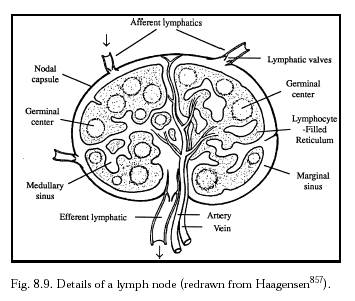
Interposed throughout the lymphatic tree are ~450 lymph nodes (Figure 8.9), lymphatic organs that serve as bacterial and particulate filters and as a final resting place for indigestible particles. Each lymph node has a blood supply representing ~0.01% of cardiac output or ~24 ml/gm-hr of blood [3138]. One in every four lymphocytes that enters a node in the blood supply exits the blood and enters the efferent lymph, and every five days ~60% of the entire blood pool of lymphocytes passes through each lymph node’s blood vessels [3138]. Despite the slow lymph velocity, filtration of lymph through the nodes can take place fairly rapidly. For example, in one experiment the subcutaneous injection of 0.15- to 0.167-micron carbon particles into murine footpads blackened regional lymph nodes in just 1-8 minutes [3129]. The very smallest particles such as ~0.05-micron liposomes generally are not retained in nodes, but larger particles such 0.5- to 0.7-micron liposomes [3139] are retained by lymph nodes [3139].
{kind=link}
Lymph node tissue is a loosely structured material consisting of a spongelike stroma and free cells in the meshes of the stroma. There are phagocytic fixed cells (lymph node histiocytes) in the sinuses that serve as filters to scavenge from the lymph, and destroy, such particles as red blood cells, bacteria, viruses, and larger dust particles imported by the respiratory tract and collected by macrophage cells of the bronchial nodes. A 5-mm lymph node probably contains a population of ~108 cells [3140], including mostly lymphocytes and both fixed histiocytes and free phagocytes. Nucleated cells in the prenodal lymph are typically 85% lymphocytes, 13% monocytes and macrophages, and 2% neutrophils (Section 8.2.1.3).
Particles injected intravascularly (e.g., intravenously) distribute rapidly to systemic lymph nodes [3141]. In one experiment [2834], colloidal carbon injected IV into mice was preferentially trapped immediately by postcapillary venules (PCV) and migrated easily out of the PCV either through the intercellular space of the PCV endothelium or by phagocytic processes as soon as 1 hour post-injection. The colloid particles were taken up by pericytes and macrophages around the PCV during the next 24 hours, conveyed to local nodes, distributed throughout the node cortex and medulla, and finally carried to the medullary lymphatic sinuses to be phagocytosed by nodal endothelial cells [2834]. Some redistribution of particles via the lymphatic sinuses from the regional lymph was observed 10-14 days after injection in different lymph nodes. In another experiment [3132], gelatinized carbon particles injected into the hepatic portal vein revealed a new pathway of liver lymphatics. Heavily carbon-laden macrophages migrated from the liver sinusoid into the interlobular connective tissue within 6 hours of the IV injection, and then entered the lymphatic vessels of the portal tract. By 9-12 hours, these macrophages began migrating into the celiac nodes via the two lymphatic pathways. From the marginal sinus in the celiac nodes, they moved into the interfollicular area of the superficial cortex, then accumulated in the paracortex after 12-24 hrs, finally ending up in the corticomedullary junction [3132]. In yet another experiment [3131], lymph-borne particles of tantalum coming from rat liver lymph entered hepatic hilar lymph nodes 7-8 hours after IV injection and were subsequently redistributed from marginal, trabecular and medullary sinuses to the paracortex (at 12-24 hours), and finally to medullary cords.
Particles injected extravascularly are distributed primarily to the lymphatic sinus, follicle and paracortex of regional lymph nodes draining from the injected sites. There is a systemic distribution of a smaller amount to distally located nodes and to liver, spleen, and bone marrow [3141]. In one study [3130], after injection of Pelikan ink into mouse footpads the macrophages that took up carbon particles in the peripheral tissue reached the regional lymph nodes via the afferent lymphatics. The particles then entered the germinal centers, mainly through the medullary pole of the lymph follicles, after migrating along their immediate exterior from their marginal sinus to their medullary pole [3130]. In another study [3142], ~0.25-micron India ink particles were subcutaneously injected into guinea pigs. A small number of fixed macrophages that were scattered throughout the germinal center (tingible body macrophages) of the popliteal lymph nodes ingested a few particles within 15-20 minutes, and there the particles remained. A larger number of itinerant ink-packed macrophages migrated from the peripheral tissues and were found preferentially in the medullary portion of the germinal center, together with many lymphoblastoid cells. The ink-packed phagocytes all exited the node after a short stay, carrying the particles away with them to another destination [3142].
Lymph nodes that become heavily overburdened with particles develop serious lesions. In one case study, a man was exposed to <0.1 mg/m3 quartz dust over a period of 30 years, leading to hilar lymph node fibrosis and calcification. Subsequently, he was exposed to 2 mg/m3 quartz dust for 5 years, which proved fatal since all alveolar-trapped dust was retained in his lungs in part because his lymph nodes were saturated with particles [3143]. A second man who had been exposed to 1.5 mg/m3 of quartz dust for 6 years experienced hilar node enlargement and subsequent calcification. When lymph nodes get overloaded with dust imported from the lungs – particularly silica, a potent lymphotrophic material – says Seal et al [3144]: “dust, accumulating in central lymph nodes, leads eventually to spread throughout the capsule and rupture into bronchi or pulmonary vessels, thereby sending dust laden activated cells back into the lungs to produce progressive massive fibrosis.”
As of 2002 there had been no studies of the behavior of diamond or sapphire particles instilled into lymph nodes, either for short-term or long-term exposures, and there had been no good studies of diamond dust inhalation risk (Section 15.1.2). Therefore at this time the possibility cannot be ruled out that indigestible diamondoid particles, accumulating in lymph nodes, might act like indigestible silica* in the same locale. However, a serious risk seems unlikely because diamond dust [652] and sapphire dust [2496] provoke no significant cytochemical reactions in macrophages, unlike quartz dust which is generally highly cytotoxic [652, 2496]. Perhaps diamond particles may act more like mica dust, which is also readily transported from lungs to lymph nodes, but once in the nodes, results only in “swollen dust-laden macrophages that retained their normal structure” for at least 1 year post-exposure [3145], with “fibrotic lesions limited to the formation of thick reticulin fibers.”
* There is some evidence that the body can slowly transport (in some as yet unknown manner) small amounts of silica, as evidenced by the occurrence of nondietary silica in human urinary calculi [3146].
Retention of microbial or other foreign particles produces swollen lymph nodes, e.g., from a normal ~0.5 cm size up to a node diameter of 0.7-1 cm in response to vaccination [3147]. But inhaled and translocated dust particles produce the most dramatic enlargements. For example, rat lungs instilled with volcanic ash produced mediastinal lymph nodes 8-18 times larger than normal “due to abundant cellular microgranuloma formation and early fibrosis” [3148]. Following an eight-day inhalation exposure in rats, silica particle-laden macrophages arrived at the posterior mediastinal lymph nodes and accumulated in granuloma-like structures without degeneration or apoptosis, increasing the weight of the lymph nodes progressively from 3.5-fold up to 35-fold at 52 weeks post-exposure [3149]. Tin particles also cause swelling in the regional draining lymph nodes [3150], and mercury produces threefold swelling in rat lymph nodes [3151].
In 2002, the maximum safe dust capacity of human lymph nodes was still unknown. However, an examination of the dust content and composition of fibrotic lung lesions and hilar lymph nodes from the lungs of British coalworkers found that the highest mean dust concentration in nodules and massive fibrosis was ~20%, mostly coal dust mixed with some fly ash which is composed of quartz, kaolin, and mica [3152]. Quartz was preferentially transported to the nodes. Dust taken from miners’ lymph nodes was 20.3% quartz but only 6.1% quartz in dust taken from the lungs [3152]. Nodes with dust-related lesions were typically 1-9 mm in diameter; only the most severe lesions were >10 mm in diameter [3153]. In a more recent study, diseased lungs from patients undergoing surgical lung resection were found to have pathological particle burdens in thoracic lymph nodes of 1010-1011 particles per gram of dry node tissue, with particles of mean diameter 0.5-0.9 microns [3096]. This implies a volumetric nodal particle burden ranging from 0.07-4%. Adopting ~1% particle burden as a reasonable maximum and assuming an initially unswollen 0.5-cm diameter lymph node, each node could safely retain ~109 particles of volume 1 micron3 per particle. Thus the entire human body could tolerate at most ~1012 of these particles (~1 cm3) if evenly distributed (the ideal case) throughout all the ~450 lymph nodes of the body. This estimate is crudely consistent with particle retention models which predict mean lung and lymph node particle burdens of 12 gm and 1.9 gm, respectively, after exposure to respirable coal mine dust at 2 mg/m2 during a 45-year working lifetime, at age 75 [5048]. (Some alternative measures of lung overload [781] suggest more conservative maximum particle burdens of only ~0.1% [781].)
The spleen could also be used for particle storage, but may be susceptible to granuloma and swelling (splenomegaly) when exposed to the same materials as those that similarly affect lymph nodes [3154], e.g., talc dust or tin particles in spleen [2969, 3155] and nodes [3150, 3156]. For instance, up to sixfold volumetric swelling has been provoked in rat spleens using IV inoculations of metallic tin particles [2969]. Assuming a ~1% maximum safe volumetric particle burden in the splenic white pulp to avoid granuloma and splenomegaly, the spleen could safely store at most another ~1 cm3 (~1012 particles) of 1-micron3 particles, producing no swelling. Despite the likely inertness of diamondoid materials, these maximum safe particle storage limits seem surprisingly low compared to likely nanomedical diagnostic and therapeutic doses. This points to the necessity for reliable phagocytic avoidance and escape protocols for medical nanorobots (Section 15.4.3.6).
Great post! Have nice day ! 🙂 tbx3w